Output from the entopeduncular nucleus, the feline equivalent of the internal segment of the globus pallidus, provides a modest direct input to the red nucleus as well as a more substantial indirect input via projections to the zona incerta and the Fields of Forel. 

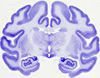
The model included the subthalamic nucleus (STN), substantia nigra (SN), zona incerta (ZI), Fields of Forel H2 (FF), internal capsule (IC) and Medtronic 3387/3389 electrode.
Additionally, stimulation through electrode contacts that improved bradykinesia and rigidity generated VTAs that overlapped the zona incerta/Fields of Forel (ZI/H2).
This suggests that HFS-STN could influence not only STN but also the dorsal adjacent structures (zona incerta and/or Fields of Forel)..
Besides the dorsolateral STN (sensorimotor part) this may include projections from/to STN, the zona incerta, and pallidofugal projections in the Fields of Forel..
The electrode on the left was within the inferior STN, whereas the right electrode was marginally superior and lateral to the intended STN target within the Fields of Forel/zona incerta. This case provides new evidence supporting cortical segregation of motor and nonmotor cortico-basal ganglionic systems that may converge in close proximity at the level of the STN and the adjacent white matter tracts (Fields of Forel/zona incerta)..
This led in 1877 to his seminal work on the organization of the tegmental region in which he provides the first description of the zona incerta and the so-called H (Haubenfeld) Fields that still bear his name.
CONCLUSIONS: Subthalamic nucleus stimulation appears to be most effective in the border area between the upper subthalamic nucleus (sensorimotor part) and the subthalamic area containing the zona incerta, Fields of Forel, and subthalamic nucleus projections..
GABA neurons of ventral thalamus (reticular nucleus, ventral lateral geniculate nucleus, zona incerta, and nucleus of the Fields of Forel) and of epithalamus appear at least 14 days before those intrinsic to dorsal thalamus.
The neurosurgical procedures used, which were guided by combined neuroimaging and neurophysiological methods, resulted in the consistent placement of DBS electrodes in the subthalamus and mesencephalon such that the electrode contacts passed through the STN and dorsally adjacent Fields of Forel (FF) and zona incerta (ZI).
Predatory ants are omnipresent year-round in upland (dryland) rice Fields in the Philippines. Solenopsis geminata flourished within many Fields, not only during the crop season but also throughout dry season fallows where they remained aggressively predatory.
The majority of cells were found contralaterally in the superior colliculus and red nucleus, and ipsilaterally in and around the interstitial nucleus of Cajal (INC), in the cuneiform region, and in the Fields of Forel.
Typical trajectories passed through the anterior thalamus, zona incerta/Fields of Forel, STN, and substantia nigra-pars reticulata.
Subcortical visual nuclei, including the ventral lateral geniculate nucleus and intergeniculate leaflet, pretectal area, and superior colliculus, and the subthalamus (zona incerta, Fields of Forel) also project to the PHA.
After raphe magnus or gigantocellular reticular pars alpha injections, a moderate to substantial number of cells were localized in the Fields of Forel, lateral habenular nucleus and ventral caudal pontine reticular nucleus.
We further revealed the presence of a substantial number of fibers in regions where glycine was not considered as a main inhibitory neurotransmitter, such as the pontine nuclei, the periaqueductal gray, the mesencephalic reticular formation, the anterior pretectal nucleus, the intralaminar thalamic nuclei, the zona incerta, the Fields of Forel, the parvocellular parts of the paraventricular nucleus, the posterior hypothalamic areas, the anterior hypothalamic area, and the lateral and medial preoptic areas.
The posterior commissure system gave rise to dense terminal Fields in the contralateral NIC, the oculomotor nucleus, and the trochlear nucleus. The descending system of NIC projections deployed dense terminal Fields in the ipsilateral gigantocellular reticular formation and the paramedian reticular formation of the pons, as well as in the ventromedial and commissural nuclei of the first two spinal cervical segments. It also gave rise to moderate or weak terminal Fields in the vestibular complex, the nucleus prepositus hypoglossi, the inferior olive, and the magnocellular reticular formation, as well as cell groups scattered along the paramedian tracts in the pons and the pontine and medullary raphe. The ascending system of NIC projections gave rise to dense terminal Fields in the ipsilateral mesencephalic reticular formation and the zona incerta as well as moderate or weak terminal Fields in the ipsilateral centromedian and parafascicular thalamic nuclei. It also provided dense bilateral labeling of the rostral interstitial nucleus of the medial longitudinal fasciculus and the Fields of Forel, and moderate or weak bilateral labeling of the mediodorsal, central medial, and central lateral nuclei of the thalamus. Models of saccade generation that rely on feedback from the velocity-to-position integrators and include the superior colliculus in their local feedback loop are contradicted because no fibers originating from the NIC traveled to the superior colliculus to deploy terminal Fields.
In addition, contralateral projections were observed to the zona incerta and the Fields of Forel.
The largest number of labelled cell bodies was found in the zona incerta, the Fields of Forel, and the ventral lateral geniculate, thalamic reticular, central medial and central lateral nuclei.
Microinjections of muscimol into the zona incerta-Forel's Fields produced tilt of the head to the injected site, while injections of bicuculline or picrotoxin produced the same effect but to the opposite side. These results imply that GABAergic mechanisms in the subthalamic nucleus and the zona incerta-Fields of Forel are involved in circling behaviour and vertical head movements, respectively..
A moderate density of dopamine-beta-hydroxylase immunoreactive processes were found in the rhomboid nucleus and zona incerta whereas little dopamine-beta-hydroxylase immunoreactivity was found in the Fields of Forel, nucleus reuniens, or subthalamic nucleus.
Targetting of the ventrolateral thalamic region was frequently accurate (77%) but because of errors in estimation of the AC-PC plane (mean forward angulation of 9 degrees) some ball bearings were placed too deeply in the Fields of and Forel, zona incerta and rostral subthalamic nucleus.
The subcortical sites were the dorsal raphe nucleus, the midbrain central gray, the Fields of Forel/zona incerta, the dorsomedial hypothalamic area, midline/intralaminar nuclei of the thalamus (posterior paraventricular, rhomboid, central medial, intermediodorsal, and mediodorsal), the medial and lateral preoptic areas, the bed nucleus of the stria terminalis, the substantia innominata, the vertical limb of the diagonal band nucleus, and the claustrum.
Disynaptic EPSPs were evoked in the motoneurones by a train of stimuli in the contralateral Fields of Forel and Zona incerta, which were due to stimulation of ascending collaterals from the tectofugal neurones. Similar disynaptic EPSPs could be evoked from the ipsilateral Fields of Forel.
These labeled structures included the central nucleus of the amygdala; the entopeduncular nucleus; the globus pallidus; the reticular and ventral lateral geniculate nuclei of the thalamus; parts of the hypothalamus including the dorsal, lateral, and posterior hypothalamic areas and the ventromedial and parvicellular nuclei; the zona incerta and Fields of Forel; parts of the substantia nigra including the pars reticularis and pars lateralis, and the retrorubral area; the pretectum; the intermediate and deep layers of the superior colliculus; the periaqueductal gray; the dorsal nucleus of the raphe; portions of the reticular formation, including the mesencephalic, pontis oralis, pontis caudalis, gigantocellularis, ventralis, and lateralis reticular nuclei; the nucleus cuneiformis; the marginal nucleus of the brachium conjunctivum; the locus coeruleus; portions of the trigeminal complex, including the principal sensory and spinal nuclei; portions of the vestibular complex, including the lateral division of the superior nucleus and the medial nucleus; deep cerebellar nuclei, including the medial and lateral cerebellar nuclei; and lamina VII of the cervical spinal cord.
A few neurons were labeled around the habenulopeduncular tract and in the medial portion of the nucleus of the Fields of Forel, in the nucleus reticularis medullaris ventralis, and in the spinal nucleus of the trigeminal nerve, the cochlear nucleus, and the superior olivary complex.
No correlation was found between the duration of aphagia and adipsia and the size of damage to zona incerta, Fields of Forel, capsula interna, lemniscus medialis, ventral tegmental area and substantia nigra.
A unilateral injection of 6-OHDA (6 microgram/1.5 microliter) was made into the Fields of Forel in order to estimate the effects of the destruction of ascending dopaminergic (DA) pathways on the denervation supersensitivity of DA D1 receptors in the rat striatum. When the destruction by 6-OHDA of this contralateral DA innervation was combined to the unilateral lesion of the Fields of Forel, the increase in DA-sensitive adenylate cyclase activity in each striatal area 3 or 7 weeks postlesion was prevented.
Transient sympathetically-mediated depressor effects were induced by stimulation of a small locus in the lateral hypothalamic peri-fornical region, medial to the Fields of Forel.
We studied nine young adult beagles at rest and during four levels of dynamic exercise before and after electrolytic lesions were made in the hypothalamus in the region of the Fields of Forel.
After large injections, 25-38% of the labeled cells were located in the brainstem reticular formation, 10-16% in the pretectal nuclei, 10-15% in the hypothalamus, 7-9% in the zona incerta, 3-9% in the Fields of Forel, 4-5% in the nucleus locus coeruleus, 3-5% in the ventral lateral geniculate body, 2-4% in the superior colliculus, 3% in the periaqueductal gray, and 14-15% in other parts of the brainstem.
Other afferents, which were thought to have been labeled through spread of HRP into the medial longitudinal fasciculus (MLF), adjacent paramedian pontine reticular formation, or uptake by transected fibers descending to the inferior olive, included the nucleus of Darkschewitsch, interstitial nucleus of Cajal, zona incerta, prerubral Fields of Forel, deep superior colliculus, nucleus of the posterior commissure, nucleus cuneiformis, ventral periaqueductal gray, vestibular complex, perihypoglossal complex, and deep cerebellar nuclei.
Furthermore, when 5-hydroxytryptophan was injected with inhibitor of monoamine oxidase, a large number of small neurons immunoreactive to serotonin was identified in many discrete regions: the anterior and lateral hypothalamic areas, preoptic area, suprachiasmatic nucleus, dorsal hypothalamic area, dorsomedial nucleus, posterior hypothalamic area and nucleus of the Fields of Forel.
Although there is considerable evidence that the H Fields of Forel of the posterior diencephalon play an important role in the regulation of cardiovascular function, little is known about the role these areas play in the control of airway caliber. In chloralose-anesthetized paralyzed dogs, we used both electrical and chemical means to stimulate the H Fields of Forel, while we monitored breath-by-breath changes in total lung resistance (TLR), a functional index of airway caliber. We conclude that stimulation of cell bodies in the H Fields of Forel produces bronchodilation by withdrawal of cholinergic tone to airway smooth muscle..
After injections of CT confined to the Mc, we found that the major afferents to the Mc arise from: (1) the lateral part of the bed nucleus of the stria terminalis, the nucleus of the anterior commissure, the preoptic area, the central nucleus of the amygdala, the posterior hypothalamus, and the nucleus of the Fields of Forel; (2) the Edinger-Westphal nucleus, the mesencephalic reticular formation, and the ventrolateral part of the periaqueductal grey; (3) the nuclei locus coeruleus alpha (LC alpha), peri-LC alpha, locus subcoeruleus, and reticularis pontis oralis and caudalis; (4) the caudal raphe nuclei; and (5) the nucleus reticularis ventralis of the medulla.(ABSTRACT TRUNCATED AT 400 WORDS).
Other sites receiving heavy projections from the RPC include adjacent regions of the pontomesencephalic reticular formation (RF), the parafascicular (PF) and central lateral (CL) nuclei of the thalamus and the Fields of Forel/zona incerta (FF-ZI).
The ventral group coursed ventrolateral from the injection site projecting to the substantia nigra, zona compacta, ventral tegmental area of Tsai, zona incerta, Fields of Forel, lateral hypothalamic nucleus and nucleus reuniens.
(1) Major rostrally oriented fibers enter the medial forebrain bundle via 3 routes which initially branch from the fasciculus retroflexus: the mediodorsal thalamic nucleus and ventromedial thalamic nucleus; the zona incerta and Fields of Forel; and the ventral tegmental area of Tsai.
Afferent sources to all of these targets were observed in the interstitial nucleus of Cajal (INC), the region surrounding the fasciculus retroflexus (PF), and the nucleus of the Fields of Forel (NFF). In the Fields of Forel, cells projecting to the vestibular nuclei or inferior olive were concentrated medially, whereas cells projecting to the spinal cord appeared both medially and laterally.
Injections involving the frontal eye Fields (FEF) labelled the INC bilaterally, but ipsilateral terminations were densest. These cases also labelled the region of the Fields of Forel.
Nucleus gigantocellularis injections produced heavy labeling in the pontomesencephalic reticular formation, the intermediate layers of the superior colliculus, the pontine and midbrain central gray, the anterior pretectal nucleus, the ventral midbrain tegmentum including the retrorubral area, the centromedian-parafascicular complex, the Fields of Forel/zona incerta, the rostral intralaminar nuclei and the lateral hypothalamic area.
Sparsely labeled sites included the Fields of Forel, substantia nigra (pars reticulata), peri-insular cortex, superior colliculus (profundum), lateral suprasylvian cortex (posterolateral lateral suprasylvian, PLLS and posteromedial lateral suprasylvian, PMLS), anterior ectosylvian cortex, thalamic reticular complex, nucleus of the optic tract, basal part of the ventromedial hypothalamic nucleus, and the pontine reticular nucleus (oralis) and adjacent reticular formation.
Following the placement of large injections of WGA-HRP into the superior colliculus of the cat, retrogradely labeled neurons are located within the reticular nucleus of the thalamus, the zona incerta, the Fields of Forel, and throughout the hypothalamus.
Results indicate that the NRTP receives afferents from visual relay nuclei, including the nucleus of optic tract, the superior colliculus, and the ventral lateral geniculate nucleus; oculomotor-associated structures including the zona incerta, the H1 and H2 Fields of Forel, the nucleus subparafasciculus, the interstitial nucleus of Cajal, the visual tegmental relay zone of the ventral tegmental area of Tsai, the mesencephalic, pontine, and medullary reticular formations, the nucleus of the posterior commissure, and a portion of the periaqueductal gray termed the supra-oculomotor periaqueductal gray; cerebellar and pontomedullary nuclei, including the superior, lateral, and medial vestibular nuclei, the deep cerebellar nuclei, and NRTP interneurons, and nuclei related to limbic functions including the lateral habenula, the mammillary nuclei, the hypothalamic nuclei, the preoptic nuclei, and the nucleus of diagonal band of Broca. The projection from the nucleus of the optic tract (NOT) confirms previous anatomical and physiological studies on the pathways involved in horizontal optokinetic nystagmus, but the number of NOT afferents is small in relation to other areas potentially related to visuomotor pathways such as the zona incerta, ventral lateral geniculate nucleus, Fields of Forel, perirubral area, and subparafasciculus.
In both monkey and cat brains, many retrogradely labeled neurons could be detected in the amygdala, hypothalamus, midline thalamus, zona incerta, and the Fields of Forel.
The Fields of Forel, the zona incerta, and lateral and dorsomedial hypothalamic areas contained a few labeled cells.
Rostral levels of the CG send heavier projections to the Fields of Forel and the zona incerta, but fewer fibers through the supraoptic decussation, than do caudal levels.
Labelled fibres and "terminal-like" labelling were found in the anterior pretectal area, in the thalamic parafascicular nucleus, in the posterior nucleus and the ventroposterior complex, in the zona incerta and in the Fields of Forel, but none were observed in the supraoptic or paraventricular nuclei. Injections made in the area of the lateral cervical nucleus and in the cuneate and gracile nuclei labelled fibres and "terminal-like" Fields in the external nucleus of the inferior colliculus, the intercollicular area, the deep grey layers of the superior colliculus and in the mesencephalic lateral tegmentum.
Most of these neurons project to the ipsilateral cortex; a small percentage innervate the contralateral cortex, but this varies among cortical terminal Fields. The perifornical neurons are organized in a roughly topographic medial-to-lateral relationship with respect to their cortical terminal Fields. Injections of different-colored fluorescent dyes into various cortical areas demonstrate that hypothalamic neurons in general have rather restricted cortical terminal Fields.
The subcortical projection from the CM-Pf complex terminates within the caudate nucleus, putamen, globus pallidus, subthalamic nucleus, zona incerta, Fields of Forel, hypothalamus, thalamic reticular nucleus, and rostral intralaminar nuclei.
Serotonergic axons were also present within the Fields of Forel and zona incerta, but they tended to avoid parts of the subthalamic nucleus.
In addition, the presence of a subthalamic locomotor region in the Fields of Forel was determined in rats after prethalamic transections..
Scant but consistent labeling occurred in the cingular, retrosplenial, and insular cortices, within the medial forebrain bundle, Fields of Forel, zona incerta, ventral tegmental area of Tsai, substantia nigra, pretectal area, periaqueductal gray, dorsal tegmental nucleus, locus ceruleus, and raphe complex.
In addition to the cerebellorubral projection, other areas of terminal labeling included the mid-brain reticular formation, nucleus parafascicularis prerubralis, zona incerta, Fields of Forel and ventral thalamus.
Dorsally, effective regions extended to the medial zona incerta and the Fields of Forel.
Following HRP injections in various parts of the inferior olive, many cells were labeled ipsilaterally in the nucleus of Darkschewitsch, the nucleus accessorius medialis of Bechterew, the nucleus of the Fields of Forel, and the subnucleus dorsomedialis and ventrolateralis of the parvocellular red nucleus. The nucleus of the Fields of Forel, suprarubral reticular formation, and interstitial nucleus of Cajal appeared to project to the caudal half of the medial accessory olive, the medial part of the ventrolateral outgrowth, the rostral part of the dorsal cap, and the caudal part of the dorsal accessory olive..
Terminal Fields were observed in laminae VII, VIII and to a lesser extent in IX. Ascending projections from NGC to the motor nucleus of V, trochlear nucleus, oculomotor nucleus, Edinger-Westphal nucleus, the ventral aspect of the periaqueductal gray, the deep and intermediate layers of the superior colliculus, nucleus parafasicularis and centromedianus, the Fields of Forel and the dorsal and lateral hypothalamic nuclei were observed. Terminal Fields are located ipsilaterally in laminae IV, V and VI, and bilaterally in VII, VIII and X.
Among the regions with few locomotion-positive sites were the subthalamic nucleus, the Fields of Forel and the habenular complex.
The results suggest that the rostral portion of the mesencephalic reticular formation receives its main input from the cerebral cortex, the zona incerta and the Fields of Forel, the central gray substance, the nuclei reticularis pontis oralis and caudalis, and the deep cerebellar nuclei.
Collaterals of these axons also terminated bilaterally in the supraoculomotor region between trochlear and oculomotor nucleus, the interstitial nucleus of Cajal and prerubral loci (including the Fields of Forel).
In contrast, fibers from the rostral pons (nucleus reticularis pontis oralis) terminate relatively heavily in the subthalamic region, including the zona incerta, the Fields of Forel, the ventral part of the thalamic reticular complex, and the lateral hypothalamus.
However, lesions placed in H1-H2 Fields of Forel and reticular formation, lateral to the periaqueductal gray, reduced the salivary response in the contralateral glands. From this study, it is concluded that the striatum is the target area for the central effect of L-DOPA on salivary secretion, by activation of pathways descending through the Fields of Forel and mesencephalic reticular formation to the contralateral lower brain stem..
In the midbrain, preterminal and terminal ramifications are present in the medial and lateral reticular tegmentum, in the central grey (including its supraoculo-motor zone), in the nuclei of Cajal and Dark-schewitsch and in the medial aspects of the prerubral area and the Fields of Forel.
These were mainly from the nucleus prepositus hypoglossi and adjacent pontine reticular formation, but also included projections from the medial vestibular an abducens nuclei and possibly subthalamic regions such as the zona incerta and Fields of Forel.
Results indicate that a significant number of afferent connections to the LH originate in the olfactory and accumbens nuclei, pyriform cortex, olfactory tracts, magnocellular and medial preoptic and anterior hypothalamic regions, stria terminalis, stria hypothalamic tract, diagonal tract of Broca, caudate-putamen and globus pallidus, internal capsule, lateral septal nuclei, lateral preoptic area and anterior medial forebrain bundle, the various amygdaloid nuclei, zona incerta, perifornical region, dorsal and ventral medial hypothalamic areas, supraoptic, paraventricular and periventricular nuclei, posterior hypothalamus and medial forebrain bundle, ventral thalamic nuclei, the Fields of Forel, arcuate and mammillary nuclei, adjacent to the fasciculus retroflexus, in the ventral tegmental area of Tsai, interpeduncular nucleus, substantia nigra, mesencephalic reticular formation, periaqueductal gray, locus coeruleus and parabrachial region.
Occasional cells were also labeled in the reticular formation lateral to the interstitial nucleus of Cajal in the caudomedial parafascicular nucleus, in the nucleus of the Fields of Forel, and in the central gray.
The hypothesis was based on previous findings that 1) psychological stress is a necessary factor for the initiation of VF in the ischemic heart of conscious pigs, 2) the frontal cortex and its related thalamic gating mechanism, uniquely show neuroelectric responses to stressful stimuli, and 3) direct electric stimulation of either the frontal cortex, posterior hypothalamus, or Fields of Forel will produce ventricular arrhythmias and myocardial necrosis. In the present study it was found that cryogenic blockade of the forebrain, posterior hypothalamus, or Fields of Forel prevents or delays VF after left anterior descending coronary artery occlusion in conscious stressed pigs (P less than 0.01).
Pontine injections of HRP also revealed that cells in Fields of Forel and zona incerta project to pons but very few subthalamic nucleus cells project there..
Bilateral stereotactic Fields of Forel surgery was carried out in 6 adults with chronic intractable epilepsy associated with multifocal EEG abnormalities. Fields of Forel surgery, while partially effective in generalized tonic clonic seizures, appears justified only in patients with such severe preexisting cognitive deficits, that the benefits of seizure control definitely outweight the risk of further cognitive decline..
In the subthalamus, the nucleus subthalamicus enlarges progressively in size, and shows a very close relationship to the zona incerta, the Fields of Forel and the pregeniculate body. The Fields of Forel is well differentiated into Fields H1 and H2.
Bilateral simultaneous stimulation elicited straight downward binocular movements from a core of tissue about 40 mm3 on each side which included the Fields of Forel, zona incerta, subthalamic nucleus, oral pole of the red nucleus, fasciculus retroflexus and 'area tegmentalis'. The minimal damage producing downward gaze defects was about 1.7 mm in diameter, cetred in the prerubral Fields, rostral and medial to the red nuclei with minimal involvement of the oral pole of these structures. These results demonstrate that the prerubral Fields contain structures which are critical for rapid eye movements downward, and therefore an isolated downward gaze palsy is a strong indicator of a bilateral lesion of this zone.
The dorsal facilitatory area appears to be an extension of the lateral division of the dorsolongitudinal fasciculus and to extend medially to join the Fields of Forel, the ventral tegmental area of Tsai, and the parvocellular, paraventricular, and periventricular nuclei.
Retrograde transport of horseradish peroxidase injected into the olive suggests that terminals within it arise from the subparafascicular nucleus of the caudal thalamus, the nucleus of Darkschewitsch, the Fields of Forel, the interstitial nucleus of Cajal, the periaqueductal grey, the caudal pretectal nucleus, the tegmentum dorsomedial to the red nucleus, the red nucleus (minimal), the nucleus linearis, as well as the dorsolateral midbrain tegmentum and tectum (Henkel et al., '75). Secondly, neurons within the subparafascicular nucleus, the nucleus of Darkschewitsch and the Fields of Forel project most heavily to parts of the medial accessory nucleus, although they also provide input to the other major subdivisions of the olive.
Three cases of intractable epilepsy treated by stereotaxic Fields of Forel Interruptions are reported with special emphasis on pre and postoperative evaluations.
In addition to its classical connection with VPLm, nucleus cuneatus projected to the following contralateral brainstem or thalamic nuclei: medial and dorsal accessory olives, external nucleus of the inferior colliculus, ventrolateral part of the superior colliculus, nucleus ruber, medial geniculate nucleus pars magnocellularis, suprageniculatus, medial and lateral divisions of the posterior thalamic nuclear group, zona incerta, and Fields of Forel.
The organization of the regional connections was outlined in a stereotaxic atlas using the autoradiographic tracing method: the majority of the ascending pathways from the rostral raphe nuclei are directed mainly through a ventrolateral bundle via the ventral tegmental area of Tsai, with some lateral extensions to the substantia nigra, and then through the Fields of Forel and the zona incerta.
More ventrally directed fibers distribute to the Fields of Forel and then spread into the posterior hypothalamus and zona incerta.
The nucleus subthalamicus and the Fields of Forel, though small in size, are comparatively well developed; the zona incerta appears to be differentiated cytoarchitectonically into two parts.
(2) Preterminal degeneration was found in: the intralaminar nuclei and the posterior group (PO); the ventral group of thalamic nuclei; the ventral thalamus, including the zona incerta, subthalamus and Fields of Forel; and the lateral hypothalamus.
from parts of the striatum to the entopeduncular nucleus (internal segment of pallidum) and from there through the H2 and H1 Fields of Forel to the oral ventral nucleus of the thalamus (V.o.a) which directly projects to the area 6 a alpha of motor cortex; 2.
-
[ View All ]